线粒体的研究是从19世纪50年代末开始的。1857年,瑞士解剖学家及生理学家阿尔伯特•冯•科立克在肌肉细胞中发现了颗粒状结构。
 |
| 线粒体 |
另外的一些科学家在其他细胞中也发现了同样的结构,证实了科立克的发现。德国病理学家及组织学家理查德•阿尔特曼将这些颗粒命名为“原生粒”(bioblast)并于1886年发明了一种鉴别这些颗粒的染色法。阿尔特曼猜测这些颗粒可能是共生于细胞内的独立生活的细菌。
1898年,德国科学家卡尔•本达因这些结构时而呈线状时而呈颗粒状,所以用希腊语中“线”和“颗粒”对应的两个词——“mitos”和“chondros”——组成“mitochondrion”来为这种结构命名,这个名称被沿用至今。一年后,美国化学家莱昂诺尔•米歇利斯开发出用具有还原性的健那绿染液为线粒体染色的方法,并推断线粒体参与某些氧化反应。这一方法于1900年公布,并由美国细胞学家埃德蒙•文森特•考德里推广。德国生物化学家奥托•海因里希•沃伯格成功完成线粒体的粗提取且分离得到一些催化与氧有关的反应的呼吸酶,并提出这些酶能被氰化物(如氢氰酸)抑制的猜想。
英国生物学家大卫•基林在1923年至1933年这十年间对线粒体内的氧化还原链(redox chain)的物质基础进行探索,辨别出反应中的电子载体——细胞色素。
线粒体的直径一般为0.5-1.0μm,长1.5-3.0μm,在光学显微镜下可见。在动物细胞中,线粒体大小受细胞代谢水平限制。不同组织在不同条件下可能产生体积异常膨大的线粒体,称为“巨线粒体”(megamitochondria):胰脏外分泌细胞中可长达10-20μm;神经元胞体中的线粒体尺寸差异很大,有的也可能长达10μm;人类成纤维细胞的线粒体则更长,可达40μm。有研究表明在低氧气分压的环境中,某些如烟草的植物的线粒体能可逆地变为巨线粒体,长度可达80μm,并形成网络。
线粒体一般呈短棒状或圆球状,但因生物种类和生理状态而异,还可呈环状、线状、哑铃状、分杈状、扁盘状或其它形状。成型蛋白(shape-forming protein)介导线粒体以不同方式与周围的细胞骨架接触或在线粒体的两层膜间形成不同的连接可能是线粒体在不同细胞中呈现出不同形态的原因。
不同生物的不同组织中线粒体数量的差异是巨大的。有许多细胞只拥有多达数千个的线粒体(如肝脏细胞中有1000-2000个线粒体),而一些细胞则只有一个线粒体(如酵母菌细胞的大型分支线粒体)。大多数哺乳动物的成熟红细胞不具有线粒体。一般来说,细胞中线粒体数量取决于该细胞的代谢水平,代谢活动越旺盛的细胞线粒体越多。
线粒体分布方向与微管一致,通常分布在细胞功能旺盛的区域:如在肾脏细胞中靠近微血管,呈平行或栅状排列;在肠表皮细胞中呈两极分布,集中在顶端和基部;在精子中分布在鞭毛中区。在卵母细胞体外培养中,随着细胞逐渐成熟,线粒体会由在细胞周边分布发展成均匀分布。线粒体在细胞质中能以微管为导轨、由马达蛋白提供动力向功能旺盛的区域迁移。
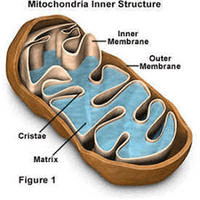
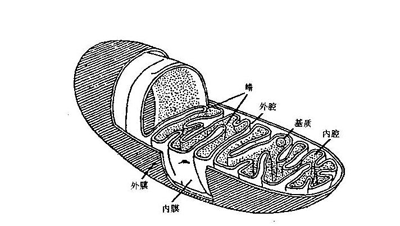
线粒体的结构
外膜
线粒体外膜是位于线粒体最外围的一层单位膜,厚度约为6-7nm。其中磷脂与蛋白质的质量为0.9:1,与真核细胞细胞膜的同一比例相近。线粒体外膜中酶的含量相对较少,其标志酶为单胺氧化酶。线粒体外膜包含称为“孔蛋白”的整合蛋白,其内部通道宽约2-3nm,这使线粒体外膜对分子量小于5000Da的分子完全通透。分子量大于上述限制的分子则需拥有一段特定的信号序列以供识别并通过外膜转运酶(translocase of the outer membrane,TOM)的主动运输来进出线粒体。
线粒体外膜主要参与诸如脂肪酸链延伸、肾上腺素氧化以及色氨酸生物降解等生化反应,它也能同时对那些将在线粒体基质中进行彻底氧化的物质先行初步分解。细胞凋亡过程中,线粒体外膜对多种存在于线粒体膜间隙中的蛋白的通透性增加,使致死性蛋白进入细胞质基质,促进了细胞凋亡。高分辨三维X射线摄影可见内质网及线粒体之间的有20%膜是紧密接触的,在这些接触位点上线粒体外膜与内质网膜通过某些蛋白质相连,形成称为“线粒体结合内质网膜”(mitochondria-associated ER-membrane,MAM)的结构。该结构在脂质的相互交换和线粒体与内质网间的钙离子信号传导等过程中都有重要作用。
 |
| 线粒体 |
膜间隙
线粒体膜间隙是线粒体外膜与线粒体内膜之间的空隙,宽约6-8nm,其中充满无定形液体。由于线粒体外膜含有孔蛋白,通透性较高,而线粒体内膜通透性较低,所以线粒体膜间隙内容物的组成与细胞质基质十分接近,含有众多生化反应底物、可溶性的酶和辅助因子等。线粒体膜间隙中还含有比细胞质基质中浓度更高的腺苷酸激酶、单磷酸激酶和二磷酸激酶等激酶,其中腺苷酸激酶是线粒体膜间隙的标志酶。线粒体膜间隙中存在的蛋白质可统称为“线粒体膜间隙蛋白质”,这些蛋白质全部在细胞质基质中合成。
内膜
线粒体内膜是位于线粒体外膜内侧、包裹着线粒体基质的单位膜。线粒体内膜中蛋白质与磷脂的质量比约为0.7:0.3,并含有大量的心磷脂(心磷脂常为细菌细胞膜的成分)。线粒体内膜的某些部分会向线粒体基质折叠形成线粒体嵴。线粒体内膜的标志酶是细胞色素氧化酶。
线粒体内膜含有比外膜更多的蛋白质(超过151种,约占线粒体所含所有蛋白质的五分之一),所以承担着更复杂的生化反应。存在于线粒体内膜中的几类蛋白质主要负责以下生理过程:特异性载体运输磷酸、谷氨酸、鸟氨酸、各种离子及核苷酸等代谢产物和中间产物;内膜转运酶(translocase of the inner membrane,TIM)运输蛋白质;参与氧化磷酸化中的氧化还原反应;参与ATP的合成;控制线粒体的分裂与融合。
嵴
线粒体嵴简称“嵴”,是线 粒体内膜向线粒体基质折褶形成的一种结构。线粒体嵴的形成增大了线粒体内膜的表面积。在不同种类的细胞中,线粒体嵴的数目、形态和排列方式可能有较大差别。线粒体嵴主要有几种排列方式,分别称为“片状嵴”(lamellar cristae)、“管状嵴”(tubular cristae)和“泡状嵴”(vesicular cristae)。片状排列的线粒体嵴主要出现在高等动物细胞的线粒体中,这些片状嵴多数垂直于线粒体长轴;管状排列的线粒体嵴则主要出现在原生动物和植物细胞的线粒体中。有研究发现,睾丸间质细胞中既存在层状嵴也存在管状嵴。线粒体嵴上有许多有柄小球体,即线粒体基粒,基粒中含有ATP合酶,能利用呼吸链产生的能量合成三磷酸腺苷。所以需要较多能量的细胞,线粒体嵴的数目一般也较多。但某些形态特殊的线粒体嵴由于没有ATP合酶,所以不能合成ATP。
基质
线粒体基质是线粒体中由线粒体内膜包裹的内部空间,其中含有参与三羧酸循环、脂肪酸氧化、氨基酸降解等生化反应的酶等众多蛋白质,所以较细胞质基质黏稠。苹果酸脱氢酶是线粒体基质的标志酶。线粒体基质中一般还含有线粒体自身的DNA(即线粒体DNA)、RNA和核糖体(即线粒体核糖体)。
线粒体DNA是线粒体中的遗传物质,呈双链环状。一个线粒体中可有一个或数个线粒体DNA分子。线粒体RNA是线粒体DNA的表达产物,RNA编辑也普遍存在于线粒体RNA中,是线粒体产生功能蛋白所必不可少的过程。线粒体核糖体是存在于线粒体基质内的一种核糖体,负责完成线粒体内进行的翻译工作。线粒体核糖体的沉降系数介干55S-56S之间。一般的线粒体核糖体由28S核糖体亚基(小亚基)和39S核糖体亚基(大亚基)组成。在这类核糖体中,rRNA约占25%,核糖体蛋白质约占75%。线粒体核糖体是已发现的蛋白质含量最高的一类核糖体。线粒体基质中存在的蛋白质统称为“线粒体基质蛋白质”,包括DNA聚合酶、RNA聚合酶、柠檬酸合成酶以及三羧酸循环酶系中的酶类。大部分线粒体基质蛋白是由核基因编码的。线粒体基质蛋白不一定只在线粒体基质中表达,它们也可以在线粒体外表达。
线粒体的功能
能量转化
 |
| 显微镜下的线粒体 |
线粒体是真核生物进行氧化代谢的部位,是糖类、脂肪和氨基酸最终氧化释放能量的场所。线粒体负责的最终氧化的共同途径是三羧酸循环与氧化磷酸化,分别对应有氧呼吸的第二、三阶段。细胞质基质中完成的糖酵解和在线粒体基质中完成的三羧酸循环在会产还原型烟酰胺腺嘌呤二核苷酸(reduced nicotinarnide adenine dinucleotide,NADH)和还原型黄素腺嘌呤二核苷酸(reduced flavin adenosine dinucleotide,FADH2)等高能分子,而氧化磷酸化这一步骤的作用则是利用这些物质还原氧气释放能量合成ATP。在有氧呼吸过程中,1分子葡萄糖经过糖酵解、三羧酸循环和氧化磷酸化将能量释放后,可产生30-32分子ATP(考虑到将NADH运入线粒体可能需消耗2分子ATP)。如果细胞所在环境缺氧,则会转而进行无氧呼吸。此时,糖酵解产生的丙酮酸便不再进入线粒体内的三羧酸循环,而是继续在细胞质基质中反应(被NADH还原成乙醇或乳酸等发酵产物),但不产生ATP。所以在无氧呼吸过程中,1分子葡萄糖只能在第一阶段产生2分子ATP。
三羧酸循环
糖酵解中生成的每分子丙酮酸会被主动运输转运穿过线粒体膜。进入线粒体基质后,丙酮酸会被氧化,并与辅酶A结合生成CO2、还原型辅酶Ⅰ和乙酰辅酶A。乙酰辅酶A是三羧酸循环(也称为“柠檬酸循环”或“Krebs循环”)的初级底物。参与该循环的酶除位于线粒体内膜的琥珀酸脱氢酶外都游离于线粒体基质中。在三羧酸循环中,每分子乙酰辅酶A被氧化的同时会产生起始电子传递链的还原型辅因子(包括3分子NADH和1分子FADH2)以及1分子三磷酸鸟苷(GTP)。
氧化磷酸化
NADH和FADH2等具有还原性的分子(在细胞质基质中的还原当量可从由逆向转运蛋白构成的苹果酸-天冬氨酸穿梭系统或通过磷酸甘油穿梭作用进入电子传递链)在电子传递链里面经过几步反应最终将氧气还原并释放能量,其中一部分能量用于生成ATP,其余则作为热能散失。在线粒体内膜上的酶复合物(NADH-泛醌还原酶、泛醌-细胞色素c还原酶、细胞色素c氧化酶)利用过程中释放的能量将质子逆浓度梯度泵入线粒体膜间隙。虽然这一过程是高效的,但仍有少量电子会过早地还原氧气,形成超氧化物等活性氧(ROS),这些物质能引起氧化应激反应使线粒体性能发生衰退。
当质子被泵入线粒体膜间隙后,线粒体内膜两侧便建立起了电化学梯度,质子就会有顺浓度梯度扩散的趋势。质子唯一的扩散通道是ATP合酶(呼吸链复合物V)。当质子通过复合物从膜间隙回到线粒体基质时,电势能被ATP合酶用于将ADP和磷酸合成ATP。这个过程被称为“化学渗透”,是一种协助扩散。彼得•米切尔就因为提出了这一假说而获得了1978年诺贝尔奖。1997年诺贝尔奖获得者保罗•博耶和约翰•瓦克阐明了ATP合酶的机制。
储存钙离子
线粒体可以储存钙离子,可以和内质网、细胞外基质等结构协同作用,从而控制细胞中的钙离子浓度的动态平衡。线粒体迅速吸收钙离子的能力使其成为细胞中钙离子的缓冲区。在线粒体内膜膜电位的驱动下,钙离子可由存在于线粒体内膜中的单向运送体输送进入线粒体基质;排出线粒体基质时则需要钠-钙交换蛋白的辅助或通过钙诱导钙释放(calcium-induced-calcium-release,CICR)机制。在钙离子释放时会引起伴随着较大膜电位变化的“钙波”(calcium wave),能激活某些第二信使系统蛋白,协调诸如突触中神经递质的释放及内分泌细胞中激素的分泌。线粒体也参与细胞凋亡时的钙离子信号转导。
其他功能
除了合成ATP为细胞提供能量等主要功能外,线粒体还承担了许多其他生理功能。
•调节膜电位并控制细胞程序性死亡:当线粒体内膜与外膜接触位点处生成了由己糖激酶(细胞质基质蛋白)、外周苯并二氮受体和电压依赖阴离子通道(线粒体外膜蛋白)、肌酸激酶(线粒体膜间隙蛋白)、ADP-ATP载体(线粒体内膜蛋白)和亲环蛋白D(线粒体基质蛋白)等多种蛋白质组成的通透性转变孔道(PT孔道)后,会使线粒体内膜通透性提高,引起线粒体跨膜电位的耗散,从而导致细胞凋亡。线粒体膜通透性增加也能使诱导凋亡因子(AIF)等分子释放进入细胞质基质,破坏细胞结构。
•细胞增殖与细胞代谢的调控;
•合成胆固醇及某些血红素。线粒体的某些功能只有在特定的组织细胞中才能展现。例如,只有肝脏细胞中的线粒体才具有对氨气(蛋白质代谢过程中产生的废物)造成的毒害解毒的功能。